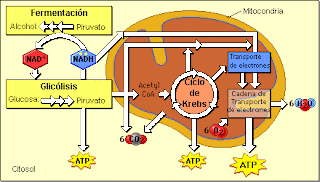
Ciclo de Krebs
También se llama ciclo de los ácidos tricarboxílicos o ciclo del citrato.
Es una vía muy importante que ocurre en la matriz mitocondrial. Es un mecanismo que incorpora el Acetil del Acetil Co-Enzima-A sobre un oxaloacetato y resulta en la eliminación de 2 C más oxidados (CO2), a expensas del paso de algunas moléculas de NAD a NADH y de FAD a FADH. Es un proceso central en el metabolismo de carbohidratos y de lípidos y de muchos aminoácidos.
Las reacciones que implica el ciclo de Krebs son:
-Una molécula de Acetil Co-A se incorpora en una molécula de oxaloacetato, dando el ácido cítrico. La condensación se da por el enzima citratosintasa. Esta reacción tiende hacia la incorporación de Acetil Co-A incluso en concentraciones muy pequeñas.
-El citrato tiene un OH central, que se transfiere a un C adyacente mediante la aconitasa. Se forma un intermedio, que es el cis-aconitato. Es una reacción de deshidratación, seguida de rehidratación. El agua que entra, entra en el otro C involucrado en el proceso.
-El isocitrato se deshidrata formando NADH por la isocitrato deshidrogenasa. Da lugar al oxalosuccinato (es químicamente inestables porque es un b-cetoácido (descarboxilan muy fácilmente) y un a-cetoácido, y pierde el carboxilo de la posición b y pasa de 6 a 5 C). Da lugar al a-cetoglutarato.
-El a-cetoglutarato, mediante la a-cetoglutarato deshidrogenasa, es descarboxilado, reduciendo un NAD, y se genera succinil co-A. Se trata de la misma reacción que realiza la piruvato deshidrogenasa. El a-cetoglutarato es un complejo proteico con 4 aminoácidos y 5 coenzimas.
-La hidrólisis del tío éster (succinil Co-A), se acopla a la síntesis de una molécula de GTP a partir de GDP. Lo hace la Succinil Co-A sintetasa). El GTP está informando a la piruvato deshidrogenasa de como va el metabolismo. El GTP pasa a ATP gracias a la nucleósido-difosfoquinasa:
-Lo que queda del ciclo, sirve para regenerar el oxaloacetato. El succinato con grupo ceto, da lugar a un oxaloacetato. Se va a oxidar este C a cetona. Se hace introduciendo 1 doble enlace entre los C centrales del succinato mediante la succinato deshidrogenasa. Se reduce FAD a FADH2. Es el único enzima que está en la membrana interna mitocondrial. Se relaciona con la cadena de transporte de electrones. Se forma el ácido fumaral.
-El fumaral es hidratado por la fumarasa, que es estereo-específica. Forma la L-Malato, que es oxidada a cetona para dar oxaloacetato mediante la malatodeshidrogenasa, que reduce NAD a NADH.
BALANCE ESTEQUIOMÉTRICO
La energía sale cuando las formas reducidas de FADH2 y NADH van a la cadena de transporte de electrones. Cada NADH rendirá 2´5 y cada FADH2, 1´5.
El ciclo de Krebs no requiere O2 en ningún lado. Sólo se necesita para regenerar FAD y NAD. Es un mecanismo anaeróbico.
Los Carbonos que entran en el ciclo de Krebs no son los mismos que salen en forma de CO2.
RELACIÓN DEL CICLO DE KREBS Y OTRAS VÍAS
El ciclo de Krebs es un punto intermedio para formar moléculas de otras vías. Es una vía anfibólicas (catabólica o anabólica).
El Acetil Co-A es también inicio de síntesis de ácidos grasos. El oxaloacetato es inicio para la síntesis de algunos aminoácidos. El a-cetoglutarato produce el esqueleto carbonado de la síntesis de algunos aminoácidos y la síntesis de purina (DNA y RNA).
El succinato es sobretodo para formar algunas porfirinas.
El fumarato se usa en la síntesis de aminoácidos y esqueletos de las pirimidinas.
La capacidad del ciclo de Krebs para regular, depende de:
-La cantidad de moléculas de oxaloacetato.
-Regulación de algún enzima.
La manera de meter Carbonos en el ciclo de Krebs es a nivel del oxaloacetato. Son 2 reacciones de relleno del ciclo de Krebs. También se llaman reacciones anapleróticas.
El paso de Acetil co-A a oxaloacetato no es posible en animales.
1. Se transforma el piruvato en oxaloacetato, catabolizado por el enzima piruvato carboxilasa. Esta reacción es muy importante, sobretodo, cuantitativamente en riñón e hígado de mamífero.
2. Del fosfoenolpiruvato (PEP), se transforma en oxaloacetato. El PEP capta CO2 y utiliza GTP en vez de ATP. Da lugar a OAA. Esta reacción puede ser reversible. Se da en músculo esquelético y cardíaco. La reacción es catalizada por la fosfoenolpiruvato carboxiquinasa (PEPCK).
Estos Carbonos pueden venir de glucosa y pueden rellenar componentes del ciclo de Krebs. Dota a la célula de un mecanismo que permite retroceder la glucólisis a través de PEP. El Piruvato puede pasar a OAA y luego a PEP invirtiendo un poco de energía.
Referencias:
(1) canal-h.net/webs/sgonzalez002/Bioquimic/GLUCOL.htm
(2) www.elergonomista.com/biologia/catabolismoglucidos.htm
(3) es.wikipedia.org/wiki/Ciclo_de_Krebs
Publicado por: Jorge José Herrera Fallas