Ciclo de la Glucólisis
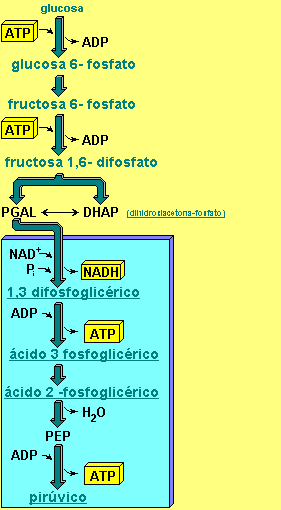
Generalidades
Durante la glucólisis se obtiene un rendimiento neto de dos moléculas de ATP y dos moléculas de NADH; el ATP puede ser usado como fuente de energía para realizar trabajo metabólico, mientras que el NADH puede tener diferentes destinos. Puede usarse como fuente de poder reductor en reacciones anabólicas; si hay oxígeno, puede oxidarse en la cadena respiratoria, obteniéndose tres ATP; si no hay oxígeno, se usa para reducir el piruvato a lactato (fermentación láctica), o a CO2 y etanol (fermentación alcohólica), sin obtención adicional de energía.Las funciones de la glucólisis son:
- La generación de moléculas de alta energía (ATP y NADH) como fuente de energía celular en procesos de respiración aeróbica (presencia de oxígeno) y fermentación (ausencia de oxígeno).
- La generación de piruvato que pasará al ciclo de Krebs, como parte de la respiración aeróbica.
- La producción de intermediarios de 6 y 3 carbonos que pueden ser utilizados en otros procesos celulares.
Descubrimiento
Los primeros estudios informales de los procesos glucolíticos fueron iniciados en 1860, cuando Louis Pasteur descubrió que los microorganismos son los responsables de la fermentación,3 y en 1897 cuando Eduard Buchner encontró que cierto extracto celular pueden causar fermentación. La siguiente gran contribución fue de Arthur Harden y William Young en 1905, quienes determinaron que para que la fermentación tenga lugar son necesarias una fracción celular de masa molecular elevada y termosensible (enzimas) y una fracción citoplasmática de baja masa molecular y termorresistente (ATP, ADP, NAD+ y otros cofactores). Los detalles de la vía en sí se determinaron en 1940, con un gran avance de Otto Meyerhoff y algunos años después por Luis Leloir. Las mayores dificultades en determinar lo intrincado de la vía fueron la corta vida y las bajas concentraciones de los intermediarios en las rápidas reacciones glicolíticas.
Reacción
- El ATP (adenosín trifosfato) es la fuente de energía universal de la célula.
- NADH y H+, otorgan la capacidad de reducir otros compuestos pertenecientes a otras vías metabólicas, o bien para sintetizar ATP.
- El piruvato es la molécula que seguirá oxidándose en el ciclo de Krebs, como parte de la respiración aeróbica, donde dará origen a más moléculas de NADH, que podrán pasar a sintetizar ATP en la mitocondria.
Destino del piruvato
Véanse también: Fermentación y Ciclo de Krebs
Luego de que una molécula de glucosa se transforme en 2 moléculas de piruvato, las condiciones del medio en que se encuentre determinarán la vía metabólica a seguir.En organismos aeróbicos, el piruvato seguirá oxidándose por la enzima piruvato deshidrogenasa y el ciclo de Krebs, creando intermediarios como NADH y FADH2. Estos intermediarios no pueden cruzar la membrana mitocondrial, y por lo tanto, utilizan sistemas de intercambio con otros compuestos llamados lanzaderas. Los más conocidos son la lanzadera malato-aspartato y la lanzadera glicerol-3-fosfato. Los intermediarios logran entregar sus equivalentes4 al interior de la membrana mitocondrial, y que luego pasarán por la cadena de transporte de electrones, que los usará para sintetizar ATP.
De esta manera, se puede obtener hasta 30 moles de ATP a partir de 1 mol de glucosa como ganancia neta.
Sin embargo, cuando las células no posean mitocondrias (Ej. eritrocito) o cuando requieran de grandes cantidades de ATP (Ej. el músculo al ejercitarse), el piruvato sufre fermentación que permite obtener 2 moles de ATP por cada mol de glucosa, por lo que esta vía es poco eficiente respecto a la fase aeróbica de la glucólisis.
Fase de gasto de energía (ATP)
Esta primera fase de la glucólisis consiste en transformar una molécula de glucosa en dos moléculas de gliceraldehído. Hasta el momento solo se ha consumido energía (ATP), sin embargo, en la segunda etapa, el gliceraldehído es convertido a una molécula de mucha energía, donde finalmente se obtendrá el beneficio final de 4 moléculas de ATP.Hexoquinasa
La primera reacción de la glucólisis es la fosforilación de la glucosa, para activarla (aumentar su energía) y así poder utilizarla en otros procesos cuando sea necesario. Esta activación ocurre por la transferencia de un grupo fosfato del ATP, una reacción catalizada por la enzima hexoquinasa,6 la cual puede fosforilar (añadir un grupo fosfato) a moléculas similares a la glucosa, como la fructosa y manosa.
Las ventajas de fosforilar la glucosa son 2: La primera es hacer de la glucosa un metabolito más reactivo, mencionado anteriormente, y la segunda ventaja es que la glucosa-6-fosfato no puede cruzar la membrana celular -a diferencia de la glucosa-ya que en la célula no existe un transportador de G6P. De esta forma se evita la pérdida de sustrato energético para la célula.
Técnicamente hablando, la hexoquinasa sólo fosforila las D-hexosas, y utiliza de sustrato Mgatp2+, ya que este catión permite que el último fosfato del ATP (fosfato gamma, γ-P o Pγ) sea un blanco más fácil para el ataque nucleofílico que realiza el grupo hidroxilo (OH) del sexto carbono de la glucosa, lo que es posible debido al Mg2+ que apantalla las cargas de los otros dos fosfatos.1 7
Fosfofructoquinasa
Fosforilación de la fructosa 6-fosfato en el carbono 1, con gasto de un ATP, a través de la enzima fosfofructoquinasa-1. También este fosfato tendrá una baja energía de hidrólisis. Por el mismo motivo que en la primera reacción, el proceso es irreversible. El nuevo producto se denominará fructosa-1,6-bifosfato.
La irreversibilidad es importante, ya que la hace ser el punto de control de la glucólisis. Como hay otros sustratos aparte de la glucosa que entran en la glucólisis, el punto de control no está colocado en la primera reacción, sino en ésta. La fosfofructoquinasa tiene centros alostéricos, sensibles a las concentraciones de intermediarios como citrato y ácidos grasos. Liberando una enzima llamada fosfructocinasa-2 que fosforila en el carbono 2 y regula la reacción.
Triosa fosfato isomerasa
Puesto que sólo el gliceraldehído-3-fosfato puede seguir los pasos restantes de la glucólisis, la otra molécula generada por la reacción anterior (dihidroxiacetona-fosfato) es isomerizada (convertida) en gliceraldehído-3-fosfato. Esta reacción posee una energía libre en condiciones estándar positiva, lo cual implicaría un proceso no favorecido, sin embargo al igual que para la reacción 4, considerando las concentraciones intracelulares reales del reactivo y el producto, se encuentra que la energía libre total es negativa, por lo que la dirección favorecida es hacia la formación de G3P.
Éste es el último paso de la "fase de gasto de energía". Sólo se ha consumido ATP en el primer paso (hexoquinasa) y el tercer paso (fosfofructoquinasa-1). Cabe recordar que el 4to paso (aldolasa) genera una molécula de gliceraldehído-3-fosfato, mientras que el 5to paso genera una segunda molécula de éste. De aquí en adelante, las reacciones a seguir ocurrirán dos veces, debido a las 2 moléculas de gliceraldehído generadas de esta fase. Hasta esta reacción hay intervención de energía (ATP).
Fase de beneficio Energético
Regulación enzimática

Gráfico que muestra la Energía libre de cada reacción en la Glucólisis
- La hexoquinasa es un punto de regulación poco importante, ya que se inhibe cuando hay mucho G-6P en músculo. Es un punto poco importante ya que el G-6P se utiliza para otras vías.
- La PFK1 es la enzima principal de la regulación de la glucólisis, actúa como una llave de agua, si está activa cataliza muchas reacciones y se obtiene más Fructosa 1,6 bifosfato, lo que permitirá a las enzimas siguientes transformar mucho piruvato. Si está inhibida, se obtienen bajas concentraciones de producto y por lo tanto se obtiene poco piruvato.
La lógica de la inhibición y activación son las siguientes:
- ATP: inhibe esta enzima pues si hay una alta concentración de ATP entonces la célula no necesita generar más.
- Citrato: Si la concentración de citrato es alta el Ciclo de Krebs va más despacio de lo que el sustrato (acetil-CoA) llega para degradarse, y la concentración de glucosa será más alta. En el Ciclo de Krebs se produce mucho NADH y FADH2, para que funcionen se han de reoxidar en la cadena de transporte electrónico creando gradiente de protones, si el gradiente no se gasta los coenzimas no se reoxidan y el Ciclo de Krebs se para.
- AMP, ADP: la alta concentración de estas moléculas implica que hay una carencia de ATP, por lo que es necesario realizar glucólisis, para generar piruvato y energía.
- La piruvato quinaza se regula distintamente según el tejido en el que trabaje, pero en hígado se inhibe en presencia de ATP y Acetil Coenzima-A (Acetil-CoA), y se activa gracias de nuevo ante la F-2,6-BP y la concentración de fosfoenolpiruvato.
Regulación por insulina
Al aumentar la glucosa en la sangre, después de una comida, las células beta del páncreas estimulan la producción de insulina, y ésta a su vez aumenta la actividad de la glucocinasa en los hepatocitos.Las concentraciones altas de glucagón y las bajas de insulina disminuyen la concentración intracelular de fructosa 2,6 bisfosfato. Esto trae por consecuencia la disminución de la glicólisis y el aumento de la gluconeogenésis.
-Lo que queda del ciclo, sirve para regenerar el oxaloacetato. El succinato con grupo ceto, da lugar a un oxaloacetato. Se va a oxidar este C a cetona. Se hace introduciendo 1 doble enlace entre los C centrales del succinato mediante la succinato deshidrogenasa. Se reduce FAD a FADH2. Es el único enzima que está en la membrana interna mitocondrial. Se relaciona con la cadena de transporte de electrones. Se forma el ácido fumaral.
-El fumaral es hidratado por la fumarasa, que es estéreo específica. Forma la L-Malato, que es oxidada a cetona para dar oxaloacetato mediante la malatodeshidrogenasa, que reduce NAD a NADH.
ANÁLISIS ESTEQUIOMÉTRICO DE LA GLUCÓLISIS
Glucosa + 2Pi + 2 ADP + 2 NAD+ 2 Pyr + 2 ATP + 2 NADH + 2 H+ + 2 H2O
La glucólisis es una vía que transforma la glucosa en Piruvato y, a su vez, reduce 2 NAD+ del citosol a NADH y usa 2 Adp para formar 2 ATP.
La célula en cuanto puede, transforma otros monosacáridos a moléculas que están en la vía de la glucólisis.
Fructosa
Se encuentra en el azúcar, se metaboliza según:
· En el hígado: la fructosa se transforma en fructosa-1-P. Se gasta 1 ATP que pasa a ADP. Lo realiza la fructoquinasa. La fructosa-1-P es degradada por una aldolasa, que da lugar a dihidroxiacetona-P y a gliceraldehido. El gliceraldehido es fosforilado con gasto de ATP a G3P. Se produce el mismo rendimiento que en la glucosa.
· En el tejido adiposo: la fructosa es fosforilada a fructosa-6-P a partir de la transformación de ATP a ADP por la hexoquinasa. La hexoquinasa tiene mucha menos afinidad por la fructosa que por la glucosa. Por eso, en el hígado, la hexoquinasa fosforila glucosa. En el tejido adiposo la hexoquinasa (HK) actúa sobre la fructosa porque no hay tanta glucosa.
La galactosa se fosforila en Gal-1-P y se gasta ATP que pasa a ADP. Lo hace la galactoquinasa. La Gal-1-P es transferida al Uridindifosfoglucosa (UDPglucosa), que es la forma activada de la glucosa, por la Gal-1-P-uridiltransferasa y se genera UDPgalactosa y se libera Glucosa-1-P Después se transforma la Glucosa-1-P en Glucosa-6-P mediante la fosfoglucomutasa.
Para que el circuito funcione correctamente, hay que regenerar UDPglucosa. Hay una epimerasa que retransforma UDP-galactosa en UDPglucosa.
Referencias:
(1) es.wikipedia.org/wiki/Glucólisis
(2) www.angelfire.com/scifi/anarkimia/Biologia/glucolisis.html
(3) www.biologia.edu.ar/metabolismo/met3glicolisis.htm
(4) canal-h.net/webs/sgonzalez002/Bioquimic/GLUCOL.htm
Publicado por: Jorge José Herrera Fallas
No hay comentarios:
Publicar un comentario