La mayoría de alumnos de bachillerato entienden por respiración celular como el simple intercambio de gases con el medio ambiente que se lleva a cabo con el aparato respiratorio y no van más allá en la cuestión, sin acabar de comprender que la verdadera respiración se lleva a cabo en cada una de nuestras células y que la función del aparato respiratorio es la de actuar como intermediario en todo el proceso de la respiración celular. Esto trae consecuencias de grandes errores conceptuales, como es el caso de: “si los vegetales carecen de aparato respiratorio, que es lo que lleva a cabo la respiración, ¡pues será que no respiran!”. (1)
Se debe entender mas bien por respiración celular que es un proceso biológico, realizado por la células vivas, en el cual, se descompone la glucosa en dióxido de carbono y agua con el fin de obtener energía química en forma de ATP contenida en las moléculas orgánicas; en otras palabras, Es claro que si todos los seres vivos necesitan utilizar energía para sus procesos metabólicos, la respiración celular por ser el medio para la liberación y utilización gradual de esa energía, se convierte en el “motor de combustión” de la célula. (2)
La respiración celular constituye el proceso más importante dentro de la célula, el cual abordaremos en pequeña medida pero de manera significativa. (3)
Esta investigación toma en cuenta a todos aquellos que de alguna manera participan aunque sea de forma mínima en la respiración celular.
Pero la respiración celular no sólo es importante por ser el medio general de obtención de energía, sino porque su rendimiento es verdaderamente excepcional. (3)
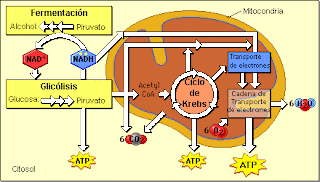
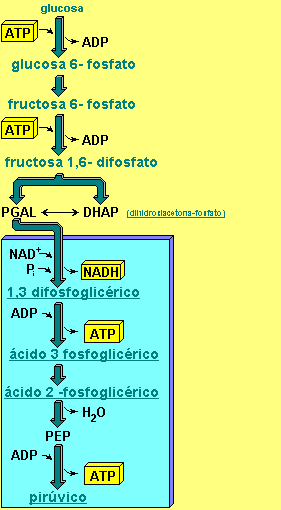
La ecuación general del proceso de respiración celular es la siguiente:
C6H12O6 (Glucosa) + 6 O2 (oxígeno) à 6 CO2 (dióxido de carbono) + 6 H2O (Agua) + ATP (energía)
Aunque esta ecuación contiene los reactivos y los productos de las reacciones generales, no describe todo el proceso; no nos dice nada de otras sustancias que son necesarias. En vez de ser una reacción simple como lo indica la reacción , la respiración consiste en una serie mas detallada de reacciones. Cada una de las reacciones que participan en el proceso está controlada por una enzima específica. Se ha calculado que en la respiración celular de la glucosa participan cerca de 100 enzimas. (2)
Esta energía en forma de ATP es aprovechada por los seres vivos en:
- Trabajo mecánico: llevado a cabo por la contracción de las fibras musculares
- Transporte activo: en el transporte celular de iones y moléculas contra sus gradientes de concentración.
- Producción de calor: para los organismos como las aves y los mamíferos que dependen del calor generado internamente (2)
“Jorge y Lucre entrenan para las competencias de atletismo de los próximos juegos intercolegiales. Durante una de las sesiones de entrenamiento Lucre sufrió un fuerte
dolor en su pierna izquierda, su entrenador manifestó que era un arratonamiento” a causa del proceso anaeróbico de fermentación láctica. (2)
Referencias:
(1) 2007. Importancia de la respiración celular. En línea, 03/08/2011. Disponible en:http://telesecundaria.setab.gob.mx/pdf/ciencias/Ciencias_Antologia06.pdf#page=79
(2) Jorge E. Rodriguez Chacón. 2008. Biología 10 y 11 año. Editograma S.A. San José, Costa Rica, Capitulo 3.5
(3) Neil A. Campbell, Jane B. Reece .2007. Biología. Mc Graw Hill. México. Capítulo 4
(4) Ministerio de Educación Pública. 2009. Programa de estudios. Educación Diversificada. Biología. San José, Costa Rica
Publicado por: Andrey Marín González